Genome-wide identification, expression profiling, and protein interaction analysis of the CCoAOMT gene family in the tea plant (Camellia sinensis)
茶树(山茶属)CCoAOMT基因家族的全基因组鉴定、表达分析和蛋白质相互作用分析,一篇全基因组家族分析的文章,有湿实验,顶配套餐了属于!各位看官有推荐评论区留言~

摘要
背景
咖啡酰辅酶A-O甲基转移酶(CCoAOMT)家族在酚类物质的氧化甲基化中起着至关重要的作用,并参与植物的多种生理过程,包括生长、发育和应对压力的反应。然而,对于茶树中CCoAOMT蛋白成员之间的相互作用了解甚少。
结果
在本研究中,我们在茶树(品种‘黄旦’)的基因组中鉴定出10个CsCCoAOMT家族成员,这些成员具有保守的基因结构和基序。这些CsCCoAOMT成员分布在六个不同的染色体(1、2、3、4、6和14)上。基于系统发育分析,CsCCoAOMT可分为两组:I 和 II。值得注意的是,Ia组的CsCCoAOMT成员可能是参与木质素生物合成的候选基因。此外,通过酵母双杂交(Y2H)实验,我们建立了CsCCoAOMT家族的蛋白相互作用网络,揭示了9对具有相互作用关系的成员。
结论
我们在茶树中鉴定了CCoAOMT基因家族,并对其分类、系统发育与同源关系、基因结构、蛋白相互作用、组织特异性表达模式和对各种压力的响应进行了全面分析。我们的发现揭示了CsCCoAOMT的进化和组成情况。特别是,观察到的CCoAOMT蛋白之间的相互作用表明,在甲基化修饰过程中可能形成甲基转移酶(OMT)复合体,这扩展了我们对该基因家族在多种生物过程中功能角色的理解。
背景
茶是全球消费量仅次于水的第二大饮料,因其众多健康益处和独特风味而受到全球消费者的喜爱。茶叶中含有多样化的次级代谢产物,包括茶多酚、嘌呤生物碱[1]和芳香化合物[2]。其中的一种芳香化合物是木质素,主要积累在次生加厚细胞中[3]。木质素为植物细胞和组织提供机械支持,帮助输送水分和养分。此外,木质素在各种生物和非生物应激反应中发挥作用[4]。然而,茶叶中木质素的积累并非总是有益的,因为它会对嫩度产生负面影响。嫩度直接影响茶叶产品的等级,并根据不同的嫩度要求采用不同的加工技术[5]。因此,茶叶中木质化的程度是评估产品等级和制定加工参数的重要参考[5]。甲基化是生物体中一种普遍且重要的化学修饰,能够改变化合物的生物活性。(-)-表儿茶素没食子酸酯(EGCG)是茶树中的主要类型儿茶素,它贡献于茶的苦味,并具有强大的抗氧化属性,带来若干健康益处[6, 7]。EGCG的另一种甲基化衍生物,(-)-表没食子儿茶素3-O-(3-O-甲基)没食子酸酯(EGCG3″Me),已被发现具有比EGCG更强的抗过敏和抗肥胖效果[8]。有趣的是,最近的研究发现,茶树(山茶属)中木质素和EGCG3"Me生物合成的相关基因属于咖啡酰辅酶A-O甲基转移酶(CCoAOMT)家族[9]。
木质素是苯丙氨酸代谢途径的重要产物,在许多植物中广泛研究,包括三种类型的单体:对羟基苯木质素(H木质素)、愈创木木质素(G木质素)和杨梅木质素(S木质素)[10]。CCoAOMT被分类为I类O-甲基转移酶(OMT),是木质素生物合成中的关键酶,负责催化咖啡酰辅酶A转化为阿魏酰辅酶A[11]。它将甲基(-CH3)从S-腺苷-L-甲硫氨酸转移到咖啡酰辅酶A的羟基(-OH)[12]。CCoAOMT在G木质素合成中起着关键作用,并为S木质素合成提供底物[13]。研究表明,高氮肥料可能会导致木质素沉积和含量降低[14]。此外,木质素的含量和组成受到生物合成途径中关键酶水平的影响,如CCoAOMT。在毛白杨中,CCoAOMT的表达也受外部氮含量的影响,不同形式和浓度的氮对PtCCoAOMT成员的表达模式产生不同的影响[15]。基因组分析结果显示,在拟南芥[12]、水稻[16]和杨树[17]中分别存在11、9和6个CCoAOMT基因。CCoAOMT包含8个保守的基序,标记为A至H。基序A(LVKVGGLIG)、B(VAPPDAPLRKY)和C(ALAVDPRIEICM)是所有OMTs中普遍存在的特征序列,而基序D(TSVYPREPEPMKELRELT)、E(KLINAKNTMEI)、F(PVIQKAGVAHKIEF)、G(DFIFVDADKDNY)和H(GDGITLCRR)是CCoAOMT特有的[18]。
在1988年,CCoAOMT在胡萝卜和欧芹的组织培养细胞中被发现,与这些组织培养细胞对真菌感染的防御反应相关[19];拟南芥中的CCoAOMT1参与抗旱应激,通过调控H2O2的积累以及ABA和ROD信号传导[20]。敲除拟南芥中编码CCoAOMT酶的基因导致G木质素含量降低,S木质素和H木质素含量增加[11, 21]。这种敲除还导致对盐胁迫的高度敏感性,通过抑制主根的生长[22]。玉米中的ZmCCoAOMT2可以调节H木质素的含量和程序性细胞死亡(PCD),在抵抗如坏死叶枯病(NLB)、南方叶枯病(SLB)、灰斑病(GLS)等疾病方面发挥重要作用[23]。最近的研究揭示,CCoAOMT不仅参与木质素的生物合成,还涉及芥子酸的代谢[11]和异黄酮素的生物合成[24],通过催化羟基肉桂酸或类黄酮前体的甲基化。此外,CCoAOMT还参与花青素的生物合成,影响植物颜色。例如,在CCoAOMT的催化下,紫荆花生成牡丹苷,导致未成熟的‘泰丽红’枣果呈紫红色[25];CCoAOMT还控制葡萄皮中花青素的甲基化过程,从而使浆果皮呈紫色[26, 27]。在茶树中,EGCG经甲基化后形成的EGCG3"Me,与EGCG相比,水溶性和生物利用度有所提高[28],从而增强了健康益处。值得注意的是,在茶树中发现CsCCoAOMT具有新的催化功能,使其能够甲基化EGCG以产生EGCG3"Me[9]。已知茶树是氟(F)的积累者,研究表明氟化物会影响茶树中儿茶素和木质素的积累,同时抑制苯丙氨酸解氨酶的活性[29]。由于CCoAOMT是儿茶素和木质素合成途径的关键调节器,它在植物多重应激响应和次级代谢产物的产生中的作用已得到确认。这凸显了它在整体植物生长和发展中的重要性。
然而,关于山茶属中CCoAOMT家族的进化关系和功能验证的研究仍然不足。本文通过生物信息学分析来弥补这一缺口,确定了‘黄旦’基因组中的CCoAOMT基因家族。此外,我们阐明了不同植物中CCoAOMT成员的进化关系,研究了CsCCoAOMT在氟和氮处理下的表达模式,并使用酵母双杂交试验检测了CsCCoAOMT成员之间的潜在相互作用。这些结果显著地增进了我们对CCoAOMT基因的理解,并为未来研究这些基因的特性提供了新的视角。
结果
茶树CCoAOMT基因家族的全基因组鉴定
通过HMM搜索,在茶树(品种‘黄旦’)的基因组中鉴定出CsCCoAOMT基因家族。最初获得了13个潜在的CsCCoAOMT基因,随后通过CDD和SMART数据库进一步分析候选基因,以去除不完整的序列。最终确定了10个CCoAOMT基因,并将它们命名为CsCCoAOMT1至CsCCoAOMT10。
CsCCoAOMT的生物信息学分析显示,鉴定的基因在开放阅读框(ORF)长度、蛋白质长度、分子量(MW)和理论等电点(pI)值方面存在变异(表S1)。ORF长度从702到864 bp不等,蛋白质长度从233氨基酸(CsCCoAOMT6)到287氨基酸(CsCCoAOMT4),分子量从26.29到32.13 kDa不等,理论等电点值从5.28到8.96不等。除了CsCCoAOMT4外,所有蛋白质均预测为稳定蛋白,且在任何蛋白中均未预测到信号肽或跨膜螺旋(TMHs)。亚细胞定位预测显示,CsCCoAOMT4和其余的CsCCoAOMT8位于叶绿体中。
CsCCoAOMT基因家族的染色体定位和同源性分析
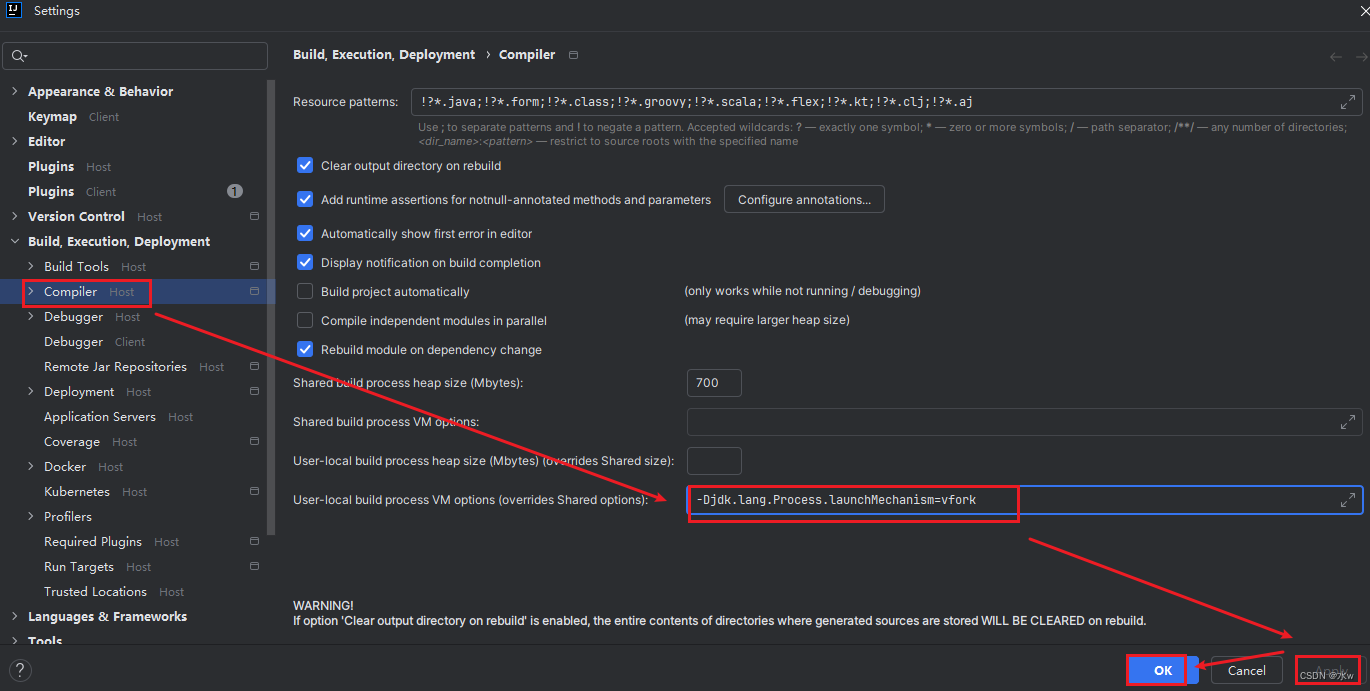
基因的染色体位置是由先前的进化事件决定的。因此,我们的调查显示,CsCCoAOMT成员随机分布在六条染色体上。其中,染色体6包含三个基因,染色体3和14各有两个基因,染色体1、2和4各有一个基因(图1A)。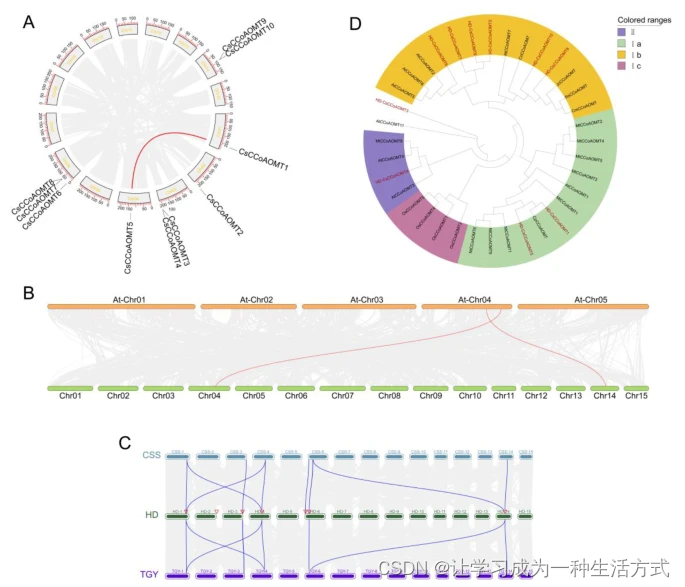
CsCCoAOMT的染色体分布、同源性分析和系统发育分析。(A) ‘黄旦’基因组中CsCCoAOMT的染色体分布和共线性分析。(B) ‘黄旦’中CsCCoAOMT与拟南芥间的物种间同源性分析。(C) ‘黄旦’、‘舒茶早’和‘铁观音’品种中CsCCoAOMT的同源性分析。(D) CsCCoAOMT与其他植物中CCoAOMT的系统发育分析。
为了阐明CsCCoAOMT基因之间的进化关系,我们分析了CsCCoAOMT家族内的同步性。我们的分析仅识别出一个同源对(CsCCoAOMT1/CsCCoAOMT5)。此外,对CsCCoAOMT的Ka/Ks比值进行了计算,CsCCoAOMT1/CsCCoAOMT5的比值为0.067,表明它们经历了净化选择。
为了进一步了解CsCCoAOMT的进化关系,我们构建了包括茶树(品种‘黄旦’)、拟南芥以及另外两个茶树品种(‘铁观音’和‘舒茶早’)的物种间比较共线性图。共线性分析显示,2个CsCCoAOMT基因与拟南芥的AtCCoAOMT表现出共线性关系(CsCCoAOMT5/AtCCoAOMT1,CsCCoAOMT10/AtCCoAOMT7)(图1B)。我们在‘黄旦’与‘舒茶早’之间发现了10对共线基因,在‘黄旦’与‘铁观音’之间发现了9对共线基因,且在‘黄旦’与‘舒茶早’之间观察到略高的同源性(图1C)。
CsCCoAOMT蛋白的系统发育分析
在拟南芥中识别了8个CCoAOMT成员,具体为AtCCoAOMT1-7和AtCCoAOMT11。序列分析显示AtCCoAOMT1-7与AtCCoAOMT11之间存在显著差异。因此,仅选择AtCCoAOMT11作为我们分析的外群[30]。
使用36个CCoAOMT蛋白构建了一个系统发育树。系统发育分析的结果显示,所有CCoAOMT蛋白被分为两个亚家族(图1D)。在进化树中,CCoAOMT蛋白被分为两个亚类:I和II。I类包括3个亚分支:Ia、Ib和Ic。Ia分支包括CsCCoAOMT1、CsCCoAOMT5和AtCCoAOMT1,这些典型的双子叶植物被证明参与木质素的生物合成。Ib亚分支由CsCCoAOMT2、CsCCoAOMT6、CsCCoAOMT7、CsCCoAOMT8、CsCCoAOMT9、CsCCoAOMT10、AtCCoAOMT2、AtCCoAOMT5、AtCCoAOMT6和AtCCoAOMT7组成。II类包括CsCCoAOMT4、AtCCoAOMT3和AtCCoAOMT4。I类与II类显示出较远的进化关系。CsCCoAOMT3与AtCCoAOMT11一起被分类为外群,表明CsCCoAOMT3与其他CCoAOMT成员有较远的进化关系。总体而言,‘黄旦’品种与拟南芥有着密切的进化关系。我们推测,茶树和拟南芥中的CCoAOMT蛋白没有明显的差异,并可能具有一些功能上的相似性。
CsCCoAOMT基因家族的基因结构和基序组成
为了更好地理解CsCCoAOMT蛋白的结构特征,我们使用MEME分析了保守基序的组成。分析预测并命名了八个保守基序,标记为基序1-8。如图2A所示,Ia分支中的所有CCoAOMT成员均具有基序1-7,而Ib分支的成员包含基序1-6,II组的成员CsCCoAOMT4含有基序1、2、5和8,而CsCCoAOMT3仅含有基序5和8(图2A)。
我们观察到CsCCoAOMT基因家族中外显子-内含子分布模式的变化,具体体现在内含子长度和外显子数量上。具体来说,CsCCoAOMT1、CsCCoAOMT5、CsCCoAOMT6、CsCCoAOMT7、CsCCoAOMT8、CsCCoAOMT9和CsCCoAOMT10具有5个外显子和4个内含子,CsCCoAOMT6含有6个外显子和5个内含子,而CsCCoAOMT4则有9个外显子和8个内含子。有趣的是,CsCCoAOMT3显示出一个独特的模式,仅有2个外显子,这使其在进化关系上远离其他成员(图2B)。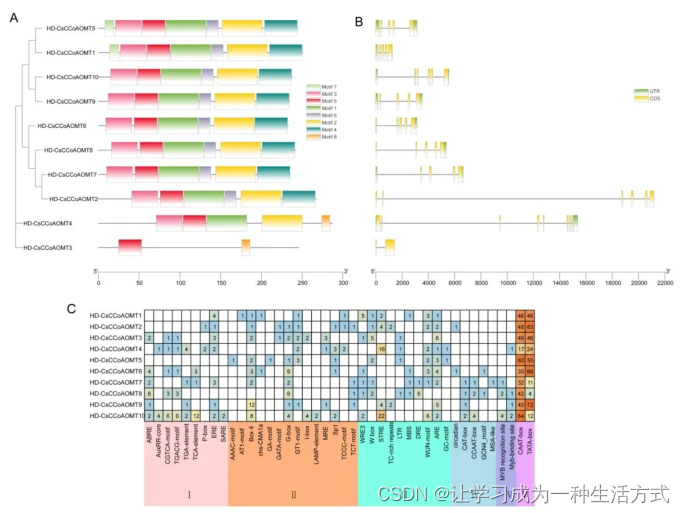
基序、外显子-内含子结构和CsCCoAOMT基因启动子中的顺式作用元件(A) CsCCoAOMTs的保守基序。(B) CsCCoAOMT的外显子-内含子结构。(C) CsCCoAOMT启动子中的顺式作用元件。注释I:植物激素响应;II:光响应;III:非生物胁迫响应;IV:植物生长;V:转录因子识别和结合位点;VI:核心。
CsCCoAOMT基因家族启动子中的顺式调控元件
我们通过提取10个CsCCoAOMT基因的上游2000个碱基序列来分析其启动子区域的顺式调控元件(图2C)。我们鉴定了33种核心启动元素,并根据它们的功能将其分为六组:光响应元件、植物激素响应元件、应激响应元件以及植物生长和发育元件。在10个CsCCoAOMT基因的上游发现了大量的光响应顺式作用元件,表明CsCCoAOMT受光信号调控。此外,还发现了各种植物激素响应元件,包括甲基茉莉酸响应元件(MeJA)、水杨酸响应元件(SA)、脱落酸响应元件(ABRE)和生长素响应元件(AUX)。CsCCoAOMT3的上游有6个ABREs,而CsCCoAOMT5有7个SA元件(图2C)。这些结果表明,CsCCoAOMT受多种植物激素调控。值得注意的是,CsCCoAOMT4、CsCCoAOMT7、CsCCoAOMT8、CsCCoAOMT9和CsCCoAOMT10基因的上游区域都包含MYB结合位点,参与了类黄酮生物合成的调控。
CsCCoAOMT的组织特异性表达模式
我们对‘黄旦’品种的8种不同组织样本中CsCCoAOMT的组织特异性表达模式进行了分析。如图3A所示,每个CsCCoAOMT基因在不同组织样本中的转录丰度有所不同。值得注意的是,CsCCoAOMT5在所有组织中表达水平较高,尤其在根和茎中。CsCCoAOMT1、CsCCoAOMT2、CsCCoAOMT3和CsCCoAOMT6在各种组织样本中的表达水平较低,而CsCCoAOMT6在果实中表达水平较高。此外,CsCCoAOMT7和CsCCoAOMT8表现出相似的表达模式,在叶片和芽中的表达水平较高。CsCCoAOMT4在成熟叶片中表达水平最高,而CsCCoAOMT9和CsCCoAOMT10在所有组织样本中均有表达。
CsCCoAOMT的表达模式。(A)不同组织中CsCCoAOMT基因的表达模式。(B)在高氮处理下,一个芽和两片叶子中CsCCoAOMT基因的表达模式。(C)在低氮处理下,一个芽和两片叶子中CsCCoAOMT基因的表达模式。(D)在氟处理下,一个芽和两片叶子中CsCCoAOMT基因的表达模式。数据被转换为log2FC(FC,折叠变化),热图用于表示CsCCoAOMT基因的响应性。蓝色和红色分别表示不同组织和处理条件下基因的下调和上调。每个点代表三个独立生物学重复的平均值。
在处理氮素水平不同时,CsCCoAOMT基因的表达模式
作为一种偏好氨的植物,茶树在土壤中对铵态氮的吸收效率很高。在本研究中,我们调查了CsCCoAOMT基因在不同氮素水平(包括高氮(HN)、低氮(LN)和氟化物(F)处理)下的表达模式。结果显示,CsCCoAOMT成员对不同外源处理的响应表达模式存在差异。在LN和HN处理下(图3B和C),CsCCoAOMT4、CsCCoAOMT5、CsCCoAOMT9和CsCCoAOMT10显示出较高的表达水平,而CsCCoAOMT1、CsCCoAOMT3和CsCCoAOMT6表现出较低的表达水平。CsCCoAOMT2、CsCCoAOMT7和CsCCoAOMT8展示了特定的表达模式,它们的表达水平在HN和LN处理后的2小时内增加,然后随着处理时间的延长(6-48小时)而下降。然而,CsCCoAOMT的表达受LN和HN处理的影响很小。
值得注意的是,茶叶中的氟含量是其他植物的数倍。在氟处理下(图3D),CsCCoAOMT2和CsCCoAOMT4的表达水平呈下降趋势。类似于外源氮处理,CsCCoAOMT1、CsCCoAOMT3和CsCCoAOMT6的表达维持在较低水平。在进行了4天的氟处理后,CsCCoAOMT9的表达达到峰值,而CsCCoAOMT7和CsCCoAOMT8在进行了2天处理后表现出上调的表达水平。
通过酵母双杂交试验检测CsCCoAOMT蛋白之间的蛋白质-蛋白质相互作用。 (A) 杂交酵母细胞在SD/-Trp-Leu-His-Ade培养基上生长。 (B) 杂交酵母细胞在含X-α-Gal的SD/-Trp-Leu-His-Ade培养基上生长。
CsCCoAOMT蛋白的相互作用
为了揭示CsCCoAOMT的蛋白质相互作用能力,我们对10个克隆成员进行了酵母双杂交分析。这10个基因的全长编码序列被插入到酵母pGBKT7载体中,以评估此系统中的自激活现象。将重组质粒和空的pGADT7载体转化到“AH109”菌株中。我们的结果显示,只有CsCCoAOMT3表现出自激活(见图S1)。
然后在这些CsCCoAOMT对之间生成了酵母杂交体(见图4)。所有酵母菌株在DDO(SD/-Trp-Leu)培养基上正常生长(见图S2)。然而,只有含有阳性二聚体的菌株能够在QDO/X(SD/-Trp-Leu-His-Ade/X-α-Gal)培养基上生长。我们的研究中鉴定了三种同源二聚体,包括CsCCoAOMT1/1、CsCCoAOMT4/4和CsCCoAOMT10/10。此外,我们还鉴定了CsCCoAOMT成员间的异源二聚体,分别是CsCCoAOMT1/5、CsCCoAOMT3/6、CsCCoAOMT3/8、CsCCoAOMT4/6、CsCCoAOMT6/8和CsCCoAOMT9/10。另外,观察到CsCCoAOMT4AD×CsCCoAOMT6BD之间的弱相互作用。总之,我们的Y2H分析鉴定了9对涉及CCoAOMT的相互作用蛋白。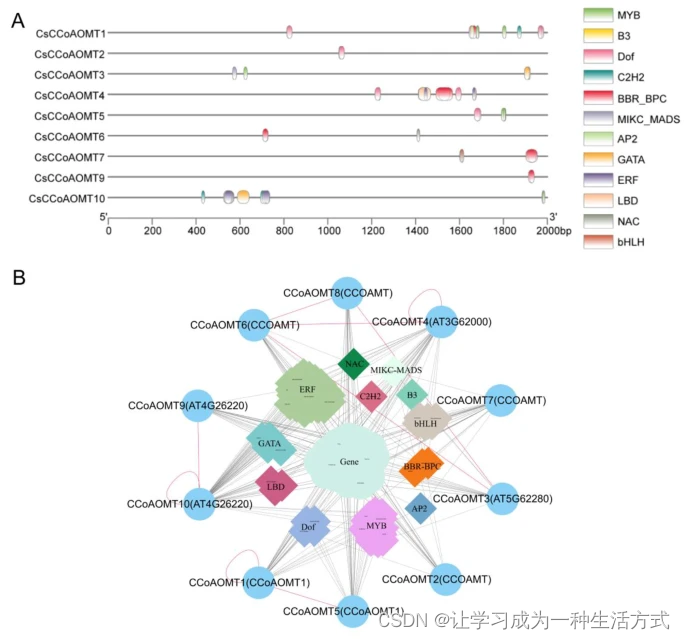
HD-CsCCoAOMT的转录因子结合位点预测及其蛋白质相互作用调控网络。(A) 在HD-CsCCoAOMT启动子中预测的转录因子结合位点。(B) HD-CsCCoAOMT的蛋白质相互作用及调控网络。最外层的圈代表‘黄旦’中的HD-CsCCoAOMT。圆圈代表功能基因,菱形代表转录因子,不同颜色的块代表不同家族的转录因子,虚线代表基因之间的相互作用,实线代表转录因子对基因的调控,红线代表初步验证。
CsCCoAOMT的相互作用与调控网络
为了研究转录因子(TF)对CsCCoAOMT表达的调控,我们使用了PlantTFDB数据库来识别启动子上的TF结合位点。我们的分析显示,有12个TF家族(MYB、B3、Dof、C2H2、BBR-BPC、MIKC-MADS、AP2、GATA、ERF、LBD、NAC和bHLH)可能与CsCCoAOMT的启动子结合,涉及的成员数量分别是16、6、12、5、12、9、3、10、51、6、1和6。在CsCCoAOMT10的启动子中识别到的TF数量和种类最多,而CsCCoAOMT8的启动子未显示任何TF结合位点(见图5A)。
我们通过结合Y2H实验、PlantTFDB数据库和STRING分析获得了CsCCoAOMT的蛋白质相互作用调控网络。具体来说,我们将十个CsCCoAOMT与拟南芥中的五个AtCCoAOMT进行了比较。预测共有27个功能基因与CsCCoAOMT发生相互作用,以及可能调控CsCCoAOMT表达的77个TF(见图5B)。值得注意的是,这些功能基因主要与木质素生物合成相关,包括4CL、HCT和C4H。
讨论
CCoAOMT是一种具有完整的AdoMet MTAses域的蛋白质,能够在植物中催化各种化合物的O-甲基化修饰。它在木质素、黄酮类和苯丙酸类化合物的生物合成中发挥关键作用。已经在不同植物种中鉴定并表征了多个CCoAOMT基因,包括拟南芥[20]、泽泻[33]和杨树[15]。然而,关于茶树中的CCoAOMT信息仍然很少[34, 35],据我们所知,尚未报道茶树中CsCCoAOMT成员之间的相互作用。
CsCCoAOMT家族的生物信息学分析
在本研究中,我们运用生物信息学方法在‘黄旦’茶树中鉴定了10个CsCCoOAMT基因,并将它们命名为CsCCoAOMT1至CsCCoAOMT10。这些基因的等电点大多低于7。我们的生物信息学分析显示,CsCCoOAMT基因在氨基酸序列、基因结构和保守基序方面显示出高度相似性。这表明CsCCoAOMT基因家族的成员虽存在一些差异,但在进化过程中保持相对保守,暗示了它们在功能上的相似性和差异性,突出了它们在调节植物生长和发育中的协同作用。
基于系统发育分析,CCoAOMT可以分为两个亚组。AtCCoAOMT1被归类为Ia分支,已确认参与拟南芥中的木质素生物合成[36]。CsCCoAOMT1和CsCCoAOMT5与AtCCoAOMT1的进化关系较近,表明它们可能参与木质素生物合成。AtCCoAOMT6被归类为Ib分支,并已显示参与拟南芥花的苯丙烷聚胺聚合物的生物合成,通过编码绒毡层O-甲基转移酶[37]。此外,体外酶学研究证实了AtCCoAOMT6对咖啡酰CoA、咖啡酰葡萄糖、绿原酸和聚胺结合物的催化活性。AtCCoAOMT7被证实在苯丙烷和黄酮生物合成中发挥作用,对黄烷酮和二氢黄烷酮的对位甲基化具有强烈的偏好[38]。我们推测,与AtCCoAOMT7同一分支的CsCCoAOMT2、CsCCoAOMT6、CsCCoAOMT7、CsCCoAOMT8、CsCCoAOMT9和CsCCoAOMT10可能参与黄酮生物合成。CsCCoAOMT3与其他CsCCoAOMT成员的进化关系较远,并与AtCCoAOMT11形成一个外群。我们推测这种蛋白可能在进化过程中发生了突变。
由于蛋白质结构决定其功能,基因结构的变异可能导致蛋白质结合构象的改变,从而显著影响基因功能。CCoAOMT的蛋白质基序相对保守,高同源性成员通常具有相似的外显子数量和保守的蛋白质基序。与系统发育树结果一致,CsCCoAOMT3的基序与其他成员差异显著,仅含有两个外显子。种间共线性分析显示,CsCCoAOMT5和CsCCoAOMT10分别与AtCCoAOMT1和AtCCoAOMT7同源。基于这些发现,我们推测这些基因可能具有相似的功能。
CsCCoAOMT家族对氮和氟处理的反应
氮对木质素和花青素的水平有显著影响。当杨树根暴露于低浓度铵氮处理(0.1 mmol/L)时,PtCCoAOMT2的表达受到抑制,而PtCCoAOMT4在下茎的表达得到促进。然而,在高浓度铵氮(10 mM)下,上茎中PtCCoAOMT1和PtCCoAOMT2的表达没有显著差异[15]。在氮限制条件下,拟南芥nla突变体中AtCCoAOMT6的表达显著上调[39]。
在我们的研究中,检测了不同氮浓度下‘黄旦’茶叶中CsCCoAOMT基因的表达水平,但基因家族成员的表达没有显著变化,表明铵含量不是影响CsCCoAOMT表达的主要因素。
茶树以其成熟叶中的高氟含量而闻名[32]。经氟处理后,CsCCoAOMT2、CsCCoAOMT4、CsCCoAOMT7和CsCCoAOMT8的表达呈现总体下降趋势,表明氟处理抑制了这些基因的表达。
CsCCoAOMT家族的相互作用关系
目前关于CCoAOMT蛋白的相互作用报道有限。例如,已证实AtCCoAOMT7能与S-腺苷-L-同型半胱氨酸水解酶(SAHH)和S-腺苷-L-甲硫氨酸合成酶(SAMS)结合,通过介导SAH降解影响细胞壁中的阿魏酸含量[40]。然而,关于茶树中CsCCoAOMT成员的相互作用蛋白没有可用信息。通过酵母双杂交实验,我们鉴定了三对同源二聚体:CsCCoAOMT1/1、CsCCoAOMT4/4和CsCCoAOMT10/10,以及五对异源二聚体:CsCCoAOMT1/5、CsCCoAOMT3/6、CsCCoAOMT3/8、CsCCoAOMT4/6和CsCCoAOMT6/8,以及CsCCoAOMT9/10。同源二聚体和异源二聚体在生物体中发挥重要的生物学作用。同源二聚体增强蛋白质的稳定性和活性,而异源二聚体通过结合不同的蛋白质
CsCCoAOMT家族的相互作用关系
目前,关于CCoAOMT蛋白的相互作用的报道非常有限。例如,已经证明AtCCoAOMT7能够与S-腺苷-L-同型半胱氨酸水解酶(SAHH)和S-腺苷-L-甲硫氨酸合成酶(SAMS)结合,通过介导SAH的降解,影响细胞壁中的阿魏酸含量[40]。然而,关于茶树中CsCCoAOMT成员的相互作用蛋白暂无信息。通过酵母双杂交实验,我们识别了三对同源二聚体:CsCCoAOMT1/1、CsCCoAOMT4/4和CsCCoAOMT10/10,以及五对异源二聚体:CsCCoAOMT1/5、CsCCoAOMT3/6、CsCCoAOMT3/8、CsCCoAOMT4/6、CsCCoAOMT6/8和CsCCoAOMT9/10。同源二聚体和异源二聚体在生物体中扮演着至关重要的生物学角色。同源二聚体增强了蛋白质的稳定性和活性,而异源二聚体通过结合不同的蛋白质单元形成新的结构和功能,从而扩展生物功能。在木薯中,MeMDH1的自我关联促进了苹果酸的生物合成,并提供疾病抵抗力。Cys330残基直接与MeMDH1的自我关联和酶活性相关[41]。桃子在TIFY成员中展示了广泛的同源二聚体和异源二聚体模式,使它们在各种生物学过程中发挥重要作用[42]。考虑到CCoAOMT在植物次生代谢产物如木质素和黄酮类的生物合成中的广泛涉及,以及它在茶树中参与EGCG3"Me的生物合成,我们推测同源和异源二聚体模式有助于增强CsCCoAOMT的酶催化活性,促进各种次生代谢产物的更高效合成,并在不同的生物学过程中发挥关键作用。
根据我们的实验数据,我们坚信CsCCoAOMT4、CsCCoAOMT5、CsCCoAOMT9和CsCCoAOMT10在茶树中扮演着重要角色。首先,qPCR结果表明这四个成员在不同组织和不同外源处理下表现出高表达趋势。此外,Y2H结果显示这四个成员与CCoAOMT的相互作用。因此,我们得出结论,CsCCoAOMT4、CsCCoAOMT5、CsCCoAOMT9和CsCCoAOMT10在山茶花中的多酚类物质的合成中发挥重要作用。
作为影响茶叶品质和等级的关键因素,新鲜叶片的嫩度与木质素含量成反比。从植物发展和抗逆生理学的角度看,木质素促进茶树的生长和发展,并增强它们的抗性[43, 44]。然而,考虑到茶饮料的质量,高木质素含量会导致叶子含量增加,影响加工和产品质量。CCoAOMT在高等植物的木质素生物合成和次生细胞壁形成中起着至关重要的作用。此外,CsCCoAOMT能够甲基化EGCG以产生O-甲基化儿茶素[9, 45],这些儿茶素与EGCG相比具有更强的抗过敏和抗肥胖效果。因此,CsCCoAOMT基因家族显著贡献于茶产品的品质和等级以及相关健康益处。
材料与方法
茶树CCoAOMT基因的鉴定
从Pfam数据库(Pfam is now hosted by InterPro)搜索并下载了CCoAOMT保守结构域(PF01596)的HMM概要文件。使用HMMER 3.0软件的搜索命令和CCoAOMT保守结构域的HMM概要文件来搜索‘黄旦’[46]蛋白文件(1e−20)(Tea Plant Information Archive(TPIA): A comprehensive knowledge database for tea plant.)。在删除冗余ID后,这些候选CCoAOMT蛋白序列通过SMART(http://smart.embl-heidelberg.de/smart/set_mode)和NCBI CDD数据库(Welcome to NCBI Batch CD-search)进行了验证,参数设置为默认。这些蛋白序列的理化性质,包括分子量(MW)、理论等电点(PI)和序列长度,由ExPASy(SIB Swiss Institute of Bioinformatics | Expasy Protein)预测。这些CsCCoAOMT基因根据染色体上的分布顺序命名。
CCoAOMT基因的系统发育树、保守基序和遗传结构分析
从NCBI(National Center for Biotechnology Information)获得了拟南芥和其他植物的CCoAOMT蛋白序列,并在表S2中显示。使用MEGA7.0软件(Home)的邻接法(NJ)构建系统发育树,引导重复次数为1000次[47]。树通过生命之树互动(iTOL: Interactive Tree Of Life)可视化。在线软件MEME(MEME - Submission form)用于从CCoAOMT蛋白序列中提取保守基序,参数选择为“选择基序数量:8”;其他参数设置为默认。系统发育树、保守基序和基因结构的综合显示由TBtools(About TBtools - CJchen's Blog)[48]进行。
染色体定位、基因重复和共线性分析
从‘黄旦’的GFF3和基因组序列文件中获取CsCCoAOMT在染色体上的位置信息。在‘黄旦’基因组以及与拟南芥和另外两种茶树品种(‘舒茶早’[49]和‘铁观音’[50])基因组的共线性分析和同源性分析是使用TBtools进行的并进行了可视化。
CCoAOMT启动子中的顺式作用调节元件
从茶树基因组序列中提取CsCCoAOMT基因上游2000 bp的启动子序列,然后提交到植物顺式作用调节元件(PlantCARE)网站(PlantCARE, a database of plant promoters and their cis-acting regulatory elements)进行顺式元素分析。
植物材料、氮和氟处理
‘黄旦’作为植物材料,购于安溪千和茶园(2020年9月)。用于组织特异性表达模式分析的样本(五年生茶苗):从茶树品种‘黄旦’收集9种组织样本(花、芽、果、嫩叶、成熟叶、老叶、嫩茎、老茎和根)。收获后,所有样本立即用液氮冷冻并保存在-80°C直到使用。
用于外源处理实验的茶苗(一年生茶苗)在水中培养。经过一个月的正常和稳定生长后,使用(NH4)2SO4作为NH4+-N来源,并包括两个处理组:高氮(9 mmol/L)和低氮(0.8 mmol/L)。处理后在0小时、2小时、6小时、24小时和48小时收集一芽两叶。我们使用NaF作为氟源来处理茶苗[51],NaF(1.2 mmol/L),并在0天、1天、2天、4天和8天收集一芽两叶。所有样本都用液氮冷冻并存放在-80°C。
每个过程收集了三个重复样本。用于基因表达分析的所有样本均进行了三次提取和分析。
总RNA提取和qRT‒PCR分析
总RNA使用天根(北京)公司的RNAprep Pure Plant Plus Kit按照厂商的说明书提取。使用TransScript® II All-in-One First-Strand cDNA Synthesis SuperMix for qPCR(Transgen, 北京, 中国)合成第一链cDNA。所有cDNA样本加入35 µl无核酸酶水并存放在−20°C直到用于qRT‒PCR分析。qRT‒PCR使用TransStart® PerfectStart TM Green qPCR SuperMix(Transgen, 北京, 中国)进行。CCoAOMT基因的特异性引物(补充表3)由Primer Premier 5.0软件设计。CsGAPDH1(KA295375.1)被选为内参基因以检测基因表达[52]。每个10 µL反应包含5 µL的2×PerfectStart Green qPCR SuperMix, 0.5 µL的正向引物, 0.5 µL的反向引物, 1 µL的cDNA和3 µL的无RNase水。扩增程序如下:95 °C持续30秒,随后进行40个循环,每个循环95 °C持续5秒和60 °C持续30秒。每次反应结束后进行熔解曲线分析以验证PCR产物的特异性。基因表达的相对量通过2−ΔΔCt方法计算(ΔCt(测试)= Ct (目标, 测试) - Ct (参考, 测试),ΔCt(校准)= Ct (目标, 校准) - Ct (参考, 校准),ΔΔCt= ΔCt(测试) - ΔCt(校准))。基因特异性引物列在表S3中。相关基因的mRNA水平的变化归一化到CsGAPDH的水平。
基因克隆和载体构建
使用‘黄旦’所有组织部分的混合样本作为材料,根据说明书提取RNA并进行逆转录。使用ACCURATE BIOTECHNOLOGY(HUNAN)CO., LTD,长沙,中国的ApexHF HS DNA Polymerase FS和特异性引物克隆CsCCoAOMT基因家族成员。通过In-Fusion方法在pGADT7和pGBKT7线性化载体上构建扩增的目标基因,并转化入大肠杆菌,筛选阳性克隆并送上桑创生物进行测序。使用Tiangen(北京,中国)的Mini Plasmid Kit从细菌溶液中提取带有正确序列的质粒,用于酵母双杂交实验[42]。
茶树CCoAOMT家族的自激活检测
pGADT7-CCoAOMT + pGBKT7和pGBKT7-CCoAOMT + pGADT7转化入酵母菌株AH109,涂布在DDO(SD/-Trp-Leu)上,在30°C培养3-5天。通过PCR检测阳性克隆。经过PCR鉴定后,它们被点样在DDO(SD/-Trp-Leu),QDO(SD/-Trp-Leu-Ade-His),和QDO/X(SD/-Trp-Leu-Ade-His + X-α-Gal)上以验证是否有自激活活性和是否可以激活报告基因。
CCoAOMT家族成员互作的酵母双杂交验证
pGADT7-CCoAOMT和pGBKT7-CCoAOMT转化入酵母菌株AH109并在30°C培养3-5天,挑选阳性克隆进行PCR检测。经过PCR鉴定后,它们被点样在QDO(SD/-Trp-Leu-Ade-His)和QDO/X(SD/-Trp-Leu-Ade-His + X-α-Gal)上以验证互作。
结论
在本研究中,我们从茶树中鉴定了10个CsCCoAOMT基因,并对其基因结构、域和保守基序进行了全面分析。我们的研究阐明了CsCCoAOMT与不同植物种类CCoAOMT之间的进化关系。通过启动子分析,我们发现CsCCoAOMT基因在光信号和激素反应中的潜在作用。此外,我们揭示了在不同组织和外源氮、氟处理下独特的基因表达模式的变化。值得注意的是,我们发现了CsCCoAOMT成员之间以前未报告的互作。总的来说,这项调查为未来的CsCCoAOMT基因特性鉴定提供了新的见解。
数据可用性
本文报告的mRNA和蛋白序列出现在TPIA(Tea Plant Information Archive(TPIA): A comprehensive knowledge database for tea plant.)数据库中,存取编号为HD.01G0032320, HD.03G0010040, HD.03012642, HD.05G0001190, HD.06G0019340, HD.03G0000700, HD.06002937, HD.03G0010060, HD.12G0019390, 和 HD.12G0019400。顺式元素从PlantCARE数据库(PlantCARE, a database of plant promoters and their cis-acting regulatory elements)获得。CsCCoAOMT家族表达数据通过qRT-PCR生成,并可根据需要从相应作者处获得。所有其他支持结果的数据都包含在文章及其附加文件中。